- [email protected]
- +91 94455 68802 |
- AP & TS: 90031 07000 |
- Other States: 74488 88954
- Toll Free : 1800 212 7669




Our Present Victory is being Celebrated.
Ranked 87th in Engineering Category by National Institutional Rankings Framework (NIRF) 2023
Transform your business idea into a successful startup through Nidhi Prayas
Vel Tech Rangarajan Dr.Sagunthala R&D Institute of Science and Technology
Value. Venture. Vanquish.

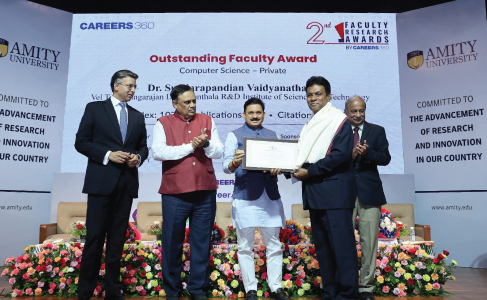
Prof. Dr. Sundarapandian Vaidyanathan from Career 360






















TAMIL NEW YEAR CELEBRATIONS
April 13, 2024

CAMPUS DRIVE SNIPE ENGINEERS
April 12, 2024


School of Computing

School of Electrical and communications

School of Mechanical and Construction

School of Sciences and Humanities

School of Management

School of Law

School of Media Technology & Communication











{kind=link}
{kind=link}